
Написано командой Qubic ScientificКоэффициент разветвления и критичность в биологических сетях, в искусственных сетях и как биовдохновленный принцип в Neuraxon
Что общего у снежной лавины, лесного пожара, землетрясения и спонтанной активности коры головного мозга?
Все они имеют границу между порядком и хаосом, что называется критическим состоянием. В мозге эта грань измеряется простым параметром: коэффициентом разветвления (σ или m). Это что-то вроде среднего соотношения "потомства" нейронов, которое каждый "родительский" нейрон активирует. Когда σ ≈ 1, активность не угасает и не взрывается; она резонирует.
Беггс и Пленц (2003) записали спонтанную активность коры головного мозга у крыс и обнаружили, что активность формировала каскадные паттерны так называемых нейронных лавин с коэффициентом ветвления, близким к 1. Казалось, что мозг находится на критической точке. У людей коэффициент ветвления σ снова оказывается близким к единице (Ван и др., 2025; Пленц и др., 2021; Вилтинг & Приесеманн, 2019).
В критической точке системы одновременно демонстрируют максимальную чувствительность к возмущениям (отзывчивость), максимальную динамическую емкость (число доступных состояний), максимальную передачу информации и максимальную сложность (Тимме и др., 2016; Шев и др., 2009, 2011).
Что такое коэффициент ветвления и как он измеряется?
Концептуально коэффициент ветвления тривиален: если в момент времени t есть A(t) активных нейронов и в t+1 есть A(t+1), тогда:
σ = ⟨ A(t+1) / A(t) ⟩
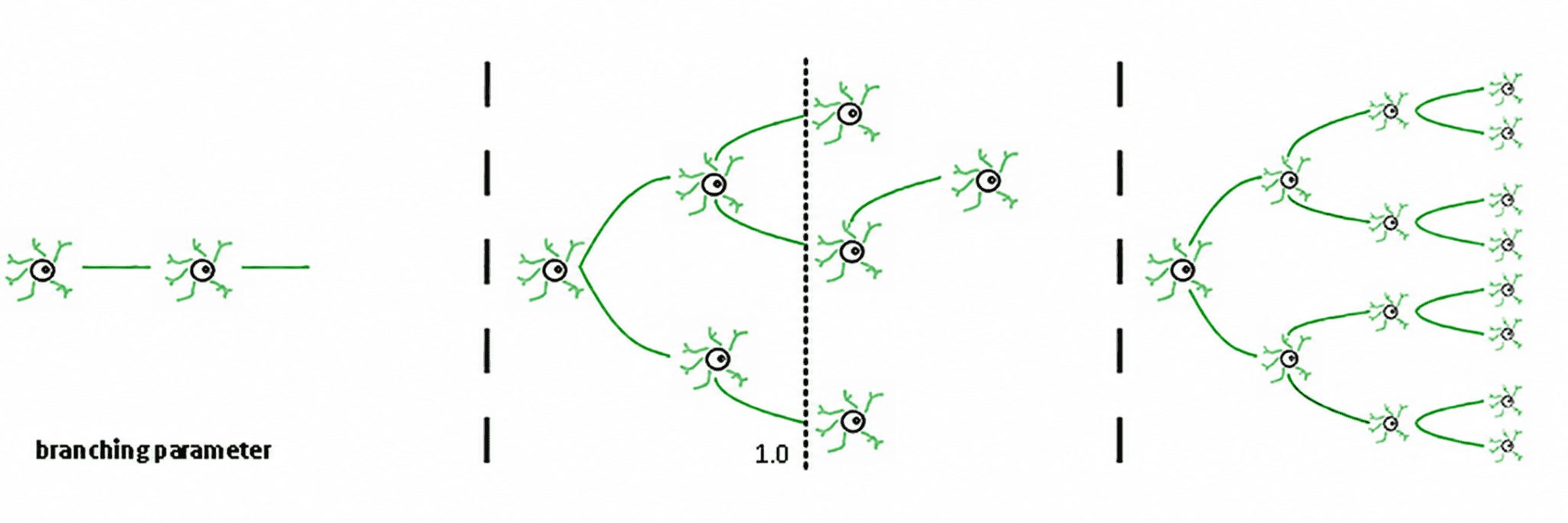
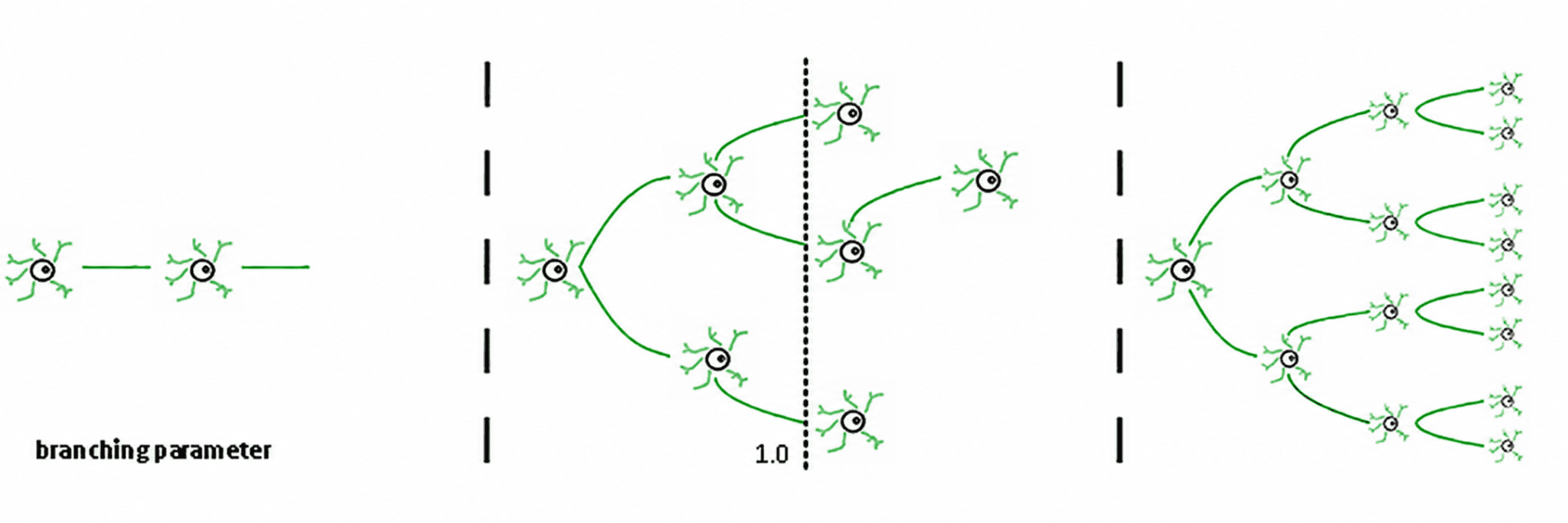
Три режима вытекают из этого (де Карвальо & Прадо, 2000; Халдеман & Беггс, 2005):
Субкритический (σ < 1): активность быстро затухает; система "забывает" возмущение. Она стабильна, но бедна в памяти и не очень выразительна.
Суперкритический (σ > 1): активность взрывается в каскады. Это характерный признак патологических режимов, таких как эпилептические припадки (Хсу и др., 2008; Хагеманн и др., 2021).
Критический (σ ≈ 1): каждая спайк, в среднем, генерирует другой спайк. Активность реверберация, нейронные лавины подчиняются степенным законам, и система поддерживает структурированную память о входных данных.
Красота σ в том, что это одно число, которое суммирует глобальный динамический режим. Но измерить его менее тривиально. При применении к in vivo корковым записям измерение показывает, что кора не работает точно при σ = 1, а немного ниже, в режиме, который авторы называют реверберационным (Вилтинг и др., 2018). Разница важна: находиться точно при σ = 1 было бы как ехать на велосипеде, балансируя на натянутом тросе; находиться немного ниже позволяет быстро настраиваться под требования задачи без риска резкого взрыва.
Критичность в искусственных нейронных сетях: от грани хаоса к резервуарным вычислениям
Бертшингер и Натшлягер (2004) показали, что случайные рекуррентные пороговые сети достигают своей максимальной вычислительной способности в задачах временной обработки именно на переходе порядка-хаоса.
Бодекер и др. (2012) расширили анализ на сети с эхостатами в рамках парадигмы резервуарных вычислений, подтвердив, что емкость передачи информации и активная память максимизируются на грани хаоса.
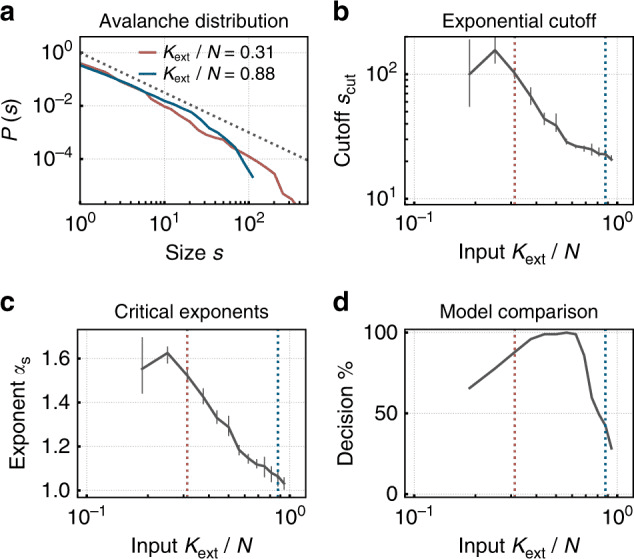
Рис. 3. Спайковая нейроморфная сеть с синаптической пластичностью самоорганизуется к критичности при низком внешнем входе, демонстрируя распределения размеров лавин с степенным законом — отличительная черта критического состояния как в биологических, так и в искусственных нейронных сетях. При более высоком входе сеть переходит в субкритический режим с усеченными распределениями. Воссоздано из Крамера и др. (2020), Природа Коммуникаций, 11, 2853. CC BY 4.0.
На языке искусственных нейронных сетей параметр измерения называется спектральным радиусом. Когда он превышает 1, траектории расходятся экспоненциально (хаос); когда он значительно ниже 1, сеть коллапсирует в фиксированную точку и теряет память. Спектральный радиус, близкий к 1, является в этом контексте формальным эквивалентом биологического σ ≈ 1 (Манагаско, 2022; Моралес и др., 2023). В спайковых нейронных сетях коэффициент ветвления можно измерить с помощью методов, почти идентичных тем, которые используются в нейронных культурах (Крамер и др., 2020; Зерати и др., 2024).
Почему критичность мозга максимизирует нейронные вычисления?
Работа вблизи σ ≈ 1 предоставляет четыре преимущества, которые являются центральными как для гипотезы критического мозга, так и для дизайна систем ИИ, вдохновленных мозгом:
Максимальный динамический диапазон. Шев и др. (2009) показали, что диапазон интенсивностей входных сигналов, которые кора может различать, максимален, когда баланс возбуждения–ингибирования помещает сеть в критичность.
Максимизированная информационная емкость. Энтропия паттернов лавин и взаимная информация между входом и выходом достигают максимума при σ ≈ 1 (Шев и др., 2011).
Оптимальная память с затуханием. В критическом режиме возмущение сохраняется ровно столько, чтобы повлиять на обработку, не загрязняя отдаленное будущее; это сладкое место между стабильностью и временной интеграцией (Бодекер и др., 2012).
Сложность как унифицированная мера. Тимме и др. (2016) продемонстрировали, что нейронная сложность максимизируется точно в критической точке, связывая критичность с формальными теориями сознания и обработки.
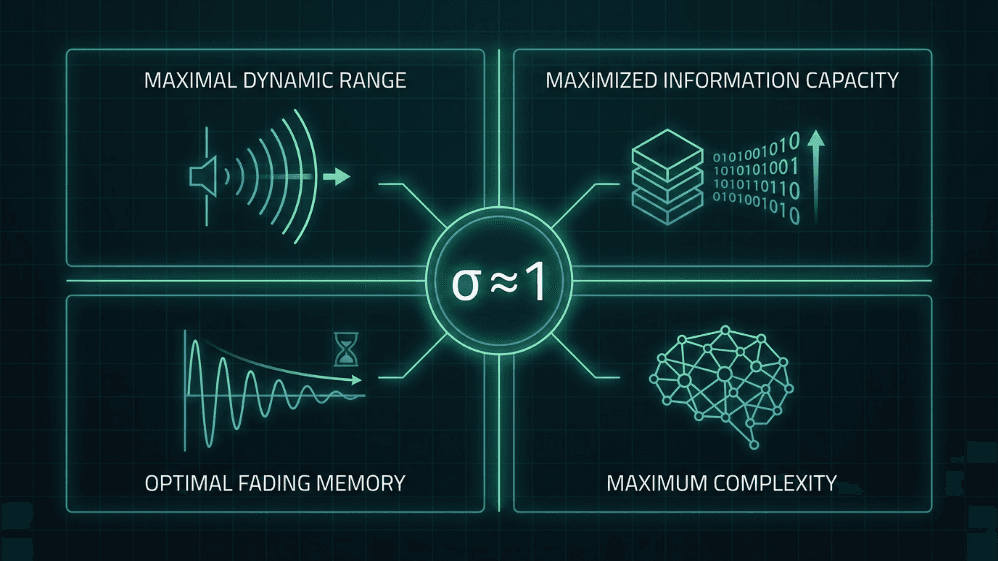
Рис. 4. Четыре вычислительных преимущества работы вблизи критического коэффициента ветвления (σ ≈ 1). При критичности нейронные сети достигают максимального динамического диапазона, максимизированной информационной емкости, оптимальной памяти с затуханием и максимальной сложности — свойства, которые являются центральными как для гипотезы критического мозга, так и для дизайна ИИ, вдохновленного мозгом.
Мозг не всегда работает при σ = 1
Это не означает, что мозг всегда работает при σ = 1. Доказательства скорее указывают на слегка субкритический и модулируемый режим: во время требовательных задач сеть приближается к критичности, во время глубокого сна она удаляется, а патологические состояния (эпилепсия, глубокая анестезия, определенные психиатрические состояния) ассоциируются с измеримыми отклонениями от этого операционного диапазона (Мейзель и др., 2017; Циммерн, 2020). Коэффициент ветвления становится динамическим биомаркером функционального состояния нервной системы.
Почему мы используем коэффициент ветвления в Neuraxon: Дизайн ИИ, вдохновленный природой, на грани хаоса
Neuraxon — это биовдохновленная система, которая принимает динамические принципы коры в качестве проектных ограничений. Коэффициент ветвления является одним из самых важных, и мы используем его по четырем причинам:
Как операционный инвариант в реальном времени для стабильности нейронной сети
В глубоких спайковых или рекуррентных архитектурах двойной риск коллапса активности (молчащая сеть, исчезающие градиенты) и бегства в взрыв (насыщение, взрывающиеся градиенты) является структурным. Мониторинг σ в реальном времени дает нам единую диагностическую скалярную величину, независимую от конкретной архитектуры, которая указывает, жива ли система в вычислительном смысле.
Как биовдохновленная цель саморегуляции через самоорганизованную критичность
Сеть самоорганизуется к критичности без необходимости централизованной тонкой настройки, воспроизводя принцип самоорганизованной критичности (Борнхольдт & Рёль, 2003; Левина и др., 2007). Это значительно снижает чувствительность к гиперпараметрам и наделяет систему устойчивостью к изменениям распределения. Как мы исследовали в NIA Том 7 о искусственной жизни и цифровых экосистемах, именно так возникает возникающая сложность из локальных правил без централизованного контроля.
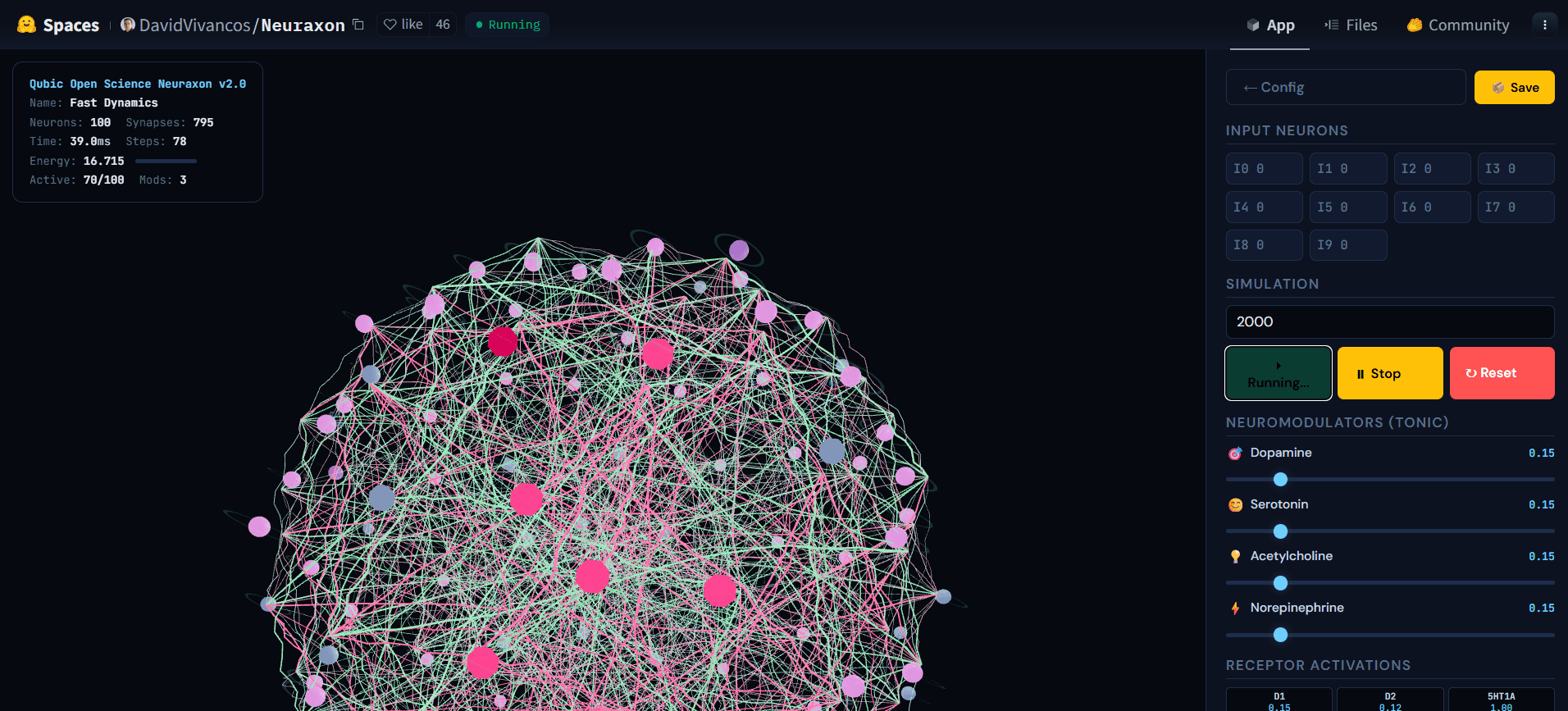
Рис. 5. 3D-сеть Neuraxon во время активной симуляции, показывающая каскадную активность среди нейронов с тройным состоянием. Ярко активные узлы (розовые) передают сигналы через возбуждающие (зеленые) и ингибирующие (розовые) соединения, в то время как другие нейроны остаются в состоянии покоя (серыми), иллюстрируя реверберационный режим рядом с критическим коэффициентом ветвления (σ ≈ 1). Это сбалансированное состояние — ни молчаливое, ни взрывное — к которому Neuraxon самоорганизуется, используя принципы критичности, вдохновленные природой. Изучите интерактивную демонстрацию на huggingface.co/spaces/DavidVivancos/Neuraxon. Источник: Научная команда Qubic.
Как мост между нейробиологическими наблюдениями и дизайном ИИ
Коэффициент ветвления — одна из очень немногих величин, которые измеряются с одинаковым формализмом в электрофизиологии, фМРТ и искусственных сетях. Это позволяет тестировать двусторонние гипотезы: если вмешательство улучшает биологическую критичность, мы можем спросить, улучшает ли то же вмешательство — переведенное в искусственную архитектуру — вычисления модели, и наоборот. Этот принцип является центральным для рамок нейромодуляции и механизмов астроцитарного гейтинга, которые мы разработали в предыдущих томах этой академии.
Как функциональный, а не эстетический критерий для ИИ, вдохновленного мозгом
Критичность является операционным ограничением с эмпирическими последствиями. Работа вблизи реверберационного режима улучшает — как измерено в наших внутренних оценках и представленных публикациях — способность к обобщению, стабильность под воздействием входных возмущений, богатство представлений и временную согласованность рассуждений. Эти эффекты качественно совпадают с теми, которые описаны как в биологической (Кокки и др., 2017), так и в искусственной (Крамер и др., 2020; Моралес и др., 2023) литературе.
Коэффициент ветвления: от статистической физики к архитектуре ИИ, вдохновленной мозгом
Коэффициент ветвления — одна из тех концептуальных редкостей: достаточно простой, чтобы свести к одной формуле, и достаточно глубокой, чтобы объединить статистическую физику, нейробиологию, ИИ и проектирование систем. Для биологического мозга σ ≈ 1 кажется режимом, в котором возникает добродетельное сочетание чувствительности, памяти, выразительности и устойчивости. Для искусственных сетей та же самая граница — переименованная как край хаоса — предсказывает максимальную вычислительную способность.
И для Neuraxon это принципиальный принцип биовдохновленного дизайна: поддающийся аудиту, саморегулирующийся и биологически значимый метрика, которая помогает нам поддерживать систему живой, в самом широком смысле этого слова.
Ссылки
Беггс, Дж. М., & Пленц, Д. (2003). Нейронные лавины в неокорковых цепях. Журнал Нейробиологии, 23(35), 11167–11177. https://doi.org/10.1523/JNEUROSCI.23-35-11167.2003
Бертшингер, Н., & Натшлягер, Т. (2004). Вычисления в реальном времени на грани хаоса в рекуррентных нейронных сетях. Нейронные вычисления, 16(7), 1413–1436. https://doi.org/10.1162/089976604323057443
Бодекер, Й., Обст, О., Лизьер, Й. Т., Мейер, Н. М., & Асада, М. (2012). Обработка информации в сетях с эхостатами на грани хаоса. Теория в бионауках, 131(3), 205–213. https://doi.org/10.1007/s12064-011-0146-8
Борнхольдт, С., & Рёль, Т. (2003). Самоорганизованные критические нейронные сети. Физический обзор E, 67(6), 066118. https://doi.org/10.1103/PhysRevE.67.066118
Кокки, Л., Голло, Л. Л., Залеский, А., & Брейкспир, М. (2017). Критичность в мозге: синтез нейробиологии, моделей и когнитивных процессов. Прогресс в нейробиологии, 158, 132–152. https://doi.org/10.1016/j.pneurobio.2017.07.002
Крамер, Б., Стёкел, Д., Крефт, М., Вибрал, М., Шеммель, Й., Мейер, К., & Приесеманн, В. (2020). Контроль критичности и вычислений в спайковых нейроморфных сетях с пластичностью. Природа Коммуникаций, 11, 2853. https://doi.org/10.1038/s41467-020-16548-3
де Карвальо, Ж. Х., & Прадо, К. П. К. (2000). Самоорганизованная критичность в модели Олами-Федера-Кристенсена. Физический обзор писем, 84(17), 4006–4009. https://doi.org/10.1103/PhysRevLett.84.4006
Деррида, Б., & Поме, Ю. (1986). Случайные сети автоматов: простая отождествленная аппроксимация. Europhysics Letters, 1(2), 45–49. https://doi.org/10.1209/0295-5075/1/2/001
Хагеманн, А., Вилтинг, Й., Самимизад, Б., Морманн, Ф., & Приесеманн, В. (2021). Оценка критичности в предприступной активности одиночных нейронов человеческой эпилептической коры. PLOS Computational Biology, 17(3), e1008773. https://doi.org/10.1371/journal.pcbi.1008773
Халдеман, К., & Беггс, Дж. М. (2005). Критическое ветвление захватывает активность в живых нейронных сетях и максимизирует число метастабильных состояний. Физический обзор писем, 94(5), 058101. https://doi.org/10.1103/PhysRevLett.94.058101
Хсу, Д., Чен, В., Хсу, М., & Беггс, Дж. М. (2008). Открытая гипотеза: эпилепсия является ли learned, и может ли она быть unlearned? Эпилепсия & Поведение, 13(3), 511–522. https://doi.org/10.1016/j.yebeh.2008.05.007
Лангтон, К. Г. (1990). Вычисление на грани хаоса: фазовые переходы и возникающее вычисление. Physica D: Нелинейные явления, 42(1–3), 12–37. https://doi.org/10.1016/0167-2789(90)90064-V
Левина, А., Херрманн, Дж. М., & Гейзель, Т. (2007). Динамические синапсы, вызывающие самоорганизованную критичность в нейронных сетях. Природа Физики, 3(12), 857–860. https://doi.org/10.1038/nphys758
Манагаско, М. О. (2022). Робустность и гибкость нейронной функции через динамическую критичность. Ээнтропия, 24(5), 591. https://doi.org/10.3390/e24050591
Мейзель, К., Клаус, А., Вязовский, В. В., & Пленц, Д. (2017). Взаимодействие между длинно- и короткосрочными временными корреляциями формирует динамику коры в разных состояниях бдительности. Журнал Нейробиологии, 37(42), 10114–10124. https://doi.org/10.1523/JNEUROSCI.0448-17.2017
Моралес, Г. Б., ди Сантос, С., & Муньос, М. А. (2023). Рассекречивание внутренней динамики биологических и искусственных нейронных сетей: от критичности к оптимальным представлениям. Фронтальные исследования в сложных системах, 1, 1276338. https://doi.org/10.3389/fcpxs.2023.1276338
Пленц, Д., Рибейро, Т. Л., Миллер, С. Р., Келлс, П. А., Вакили, А., & Чапек, Е. Л. (2021). Самоорганизованная критичность в мозге. Фронтальные исследования в физике, 9, 639389. https://doi.org/10.3389/fphy.2021.639389
Шев, В. Л., Янг, Х., Петерманн, Т., Рой, Р., & Пленц, Д. (2009). Нейронные лавины подразумевают максимальный динамический диапазон в корковых сетях при критичности. Журнал Нейробиологии, 29(49), 15595–15600. https://doi.org/10.1523/JNEUROSCI.3864-09.2009
Шев, В. Л., Янг, Х., Ю, С., Рой, Р., & Пленц, Д. (2011). Информационная емкость и передача максимизируются в сбалансированных корковых сетях с нейронными лавинами. Журнал Нейробиологии, 31(1), 55–63. https://doi.org/10.1523/JNEUROSCI.4637-10.2011
Шпицнер, Ф. П., Дехнинг, Й., Вилтинг, Й., Хагеманн, А., Нето, Ж. П., Зиренберг, Й., & Приесеманн, В. (2021). MR. Estimator, инструмент для определения внутренних временных шкал на основе субвыборок спайковой активности. PLOS ONE, 16(4), e0249447. https://doi.org/10.1371/journal.pone.0249447
Тимме, Н. М., Маршалл, Н. Дж., Беннет, Н., Рипп, М., Лаутценхайзер, Е., & Беггс, Дж. М. (2016). Критичность максимизирует сложность в нейронной ткани. Фронтальные исследования в физиологии, 7, 425. https://doi.org/10.3389/fphys.2016.00425
Турригиано, Г. Г. (2008). Самонастраивающийся нейрон: синаптическое масштабирование возбуждающих синапсов. Клетка, 135(3), 422–435. https://doi.org/10.1016/j.cell.2008.10.008
Ван, Дж., Цао, Р., Брунтон, Б. В., Смит, Р. Е. В., Бакнер, Р. Л., & Лю, Т. Т. (2025). Генетические вклады в мозговую критичность и ее связь с когнитивными функциями человека. Труды Национальной академии наук, 122(26), e2417010122. https://doi.org/10.1073/pnas.2417010122
Вилтинг, Й., Дехнинг, Й., Пинейро Нето, Ж., Рудельт, Л., Вибрал, М., Зиренберг, Й., & Приесеманн, В. (2018). Работа в режиме реверберации позволяет быстро настраивать состояния сети под требования задачи. Фронтальные исследования в системной нейробиологии, 12, 55. https://doi.org/10.3389/fnsys.2018.00055
Вилтинг, Й., & Приесеманн, В. (2018). Выявление коллективных динамических состояний из широко не наблюдаемых систем. Природа Коммуникации, 9, 2325. https://doi.org/10.1038/s41467-018-04725-4
Вилтинг, Й., & Приесеманн, В. (2019). 25 лет критичности в нейробиологии — Установленные результаты, открытые споры, новые концепции. Текущие мнения в нейробиологии, 58, 105–111. https://doi.org/10.1016/j.conb.2019.08.002
Ю, Ц. (2022). К единой аналитике гипотезы критичности мозга: обзор нескольких доступных инструментов. Фронтальные исследования в нейронных цепях, 16, 911245. https://doi.org/10.3389/fncir.2022.911245
Зерати, Р., Энгель, Т. А., & Левина, А. (2024). Оценка внутренних временных шкал и критичности по нейронным записям: методы и подводные камни. Текущие мнения в нейробиологии, 86, 102871. https://doi.org/10.1016/j.conb.2024.102871
Циммерн, В. (2020). Почему критичность мозга имеет клиническое значение: обзор. Фронтальные исследования в нейронных цепях, 14, 54. https://doi.org/10.3389/fncir.2020.00054
Изучите Полную Академию Интеллекта Neuraxon
Это Том 8 #Neuraxon Интеллекта #academy от #Qubic Научной команды. Если вы только что присоединились, изучите всю серию, чтобы получить полное представление о науке, стоящей за Neuraxon, #aigarth и подходом Qubic к мозгоподобному, #decentralized искусственному интеллекту:
NIA Vol. 1: Почему интеллект не вычисляется шаг за шагом, а во времени — исследует, почему биологический интеллект работает в непрерывном времени, а не в дискретных вычислительных шагах, как традиционные LLM.
NIA Vol. 2: Третичные динамики как модель живого интеллекта — объясняет третичные динамики и почему логика трех состояний (возбуждающая, нейтральная, ингибирующая) важна для моделирования живых систем.
NIA Vol. 3: Нейромодуляция и искусственный интеллект, вдохновленный мозгом — охватывает нейромодуляцию и как химическая сигнализация мозга (дофамин, серотонин, ацетилхолин, норэпинефрин) вдохновляет архитектуру Neuraxon.
NIA Vol. 4: Нейронные сети в ИИ и нейробиологии — глубокое сравнение биологических нейронных сетей, искусственных нейронных сетей и третьего подхода Neuraxon.
NIA Vol. 5: Астроглии и искусственный интеллект, вдохновленный мозгом — как астроцитарные механизмы управления формируют пластичность нейронной сети через рамки AGMP в Neuraxon.
NIA Vol. 6: Сознательные машины против интеллектуальных организмов: объяснение сознания ИИ — исследует сознание ИИ через призму теории глобального рабочего пространства, теории интегрированной информации и предсказательного кодирования.
NIA Vol. 7: Игра жизни Конвея, искусственная жизнь и цифровые экосистемы — наука за Qubic, Aigarth и подход Neuraxon к возникающей сложности и самоорганизованной критичности в децентрализованном ИИ.
Qubic — это децентрализованная, открытая сеть для экспериментальных технологий. Чтобы узнать больше, посетите qubic.org. Присоединяйтесь к обсуждению на X, Discord и Telegram.