
Escrito por el equipo de Qubic ScientificLa razón de ramificación y la criticalidad en redes biológicas, en redes artificiales, y como un principio bioinspirado en Neuraxon
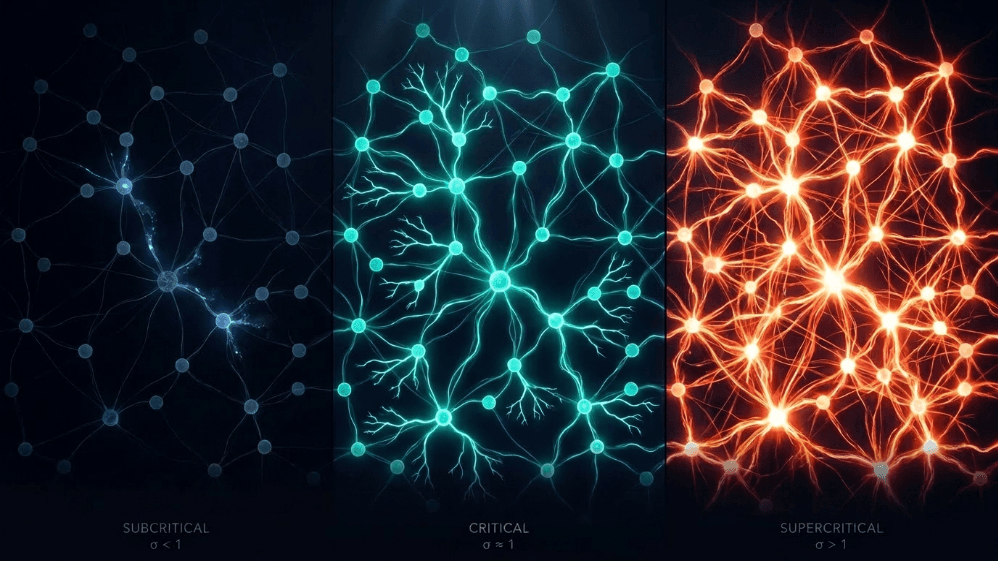
¿Qué tienen en común una avalancha de nieve, un incendio forestal, un terremoto y la actividad espontánea de la corteza cerebral?
Todos comparten una frontera entre el orden y el caos, lo que se llama un estado crítico. En el cerebro, ese límite se mide por un parámetro simple: la razón de ramificación (σ o m). Sería algo así como la razón promedio de "descendientes" neuronales que cada neurona "padre" activa. Cuando σ ≈ 1, la actividad ni se apaga ni explota; reverbera.
Beggs y Plenz (2003) registraron la actividad espontánea de la corteza cerebral en ratas y descubrieron que esta actividad formaba patrones en cascada, las llamadas avalanchas neuronales, con una relación de ramificación cercana a 1. El cerebro parecía vivir en un punto crítico. En humanos, la relación de ramificación σ vuelve a ser cercana a la unidad (Wang et al., 2025; Plenz et al., 2021; Wilting y Priesemann, 2019).
En el punto crítico, los sistemas exhiben simultáneamente máxima sensibilidad a las perturbaciones (capacidad de respuesta), máxima capacidad dinámica (número de estados accesibles), máxima transmisión de información y máxima complejidad (Timme et al., 2016; Shew et al., 2009, 2011).
¿Qué es la relación de ramificación y cómo se mide?
Conceptualmente, la relación de ramificación es trivial: si en el instante t hay A(t) neuronas activas y en t+1 hay A(t+1), entonces:
σ = ⟨ A(t+1) / A(t) ⟩
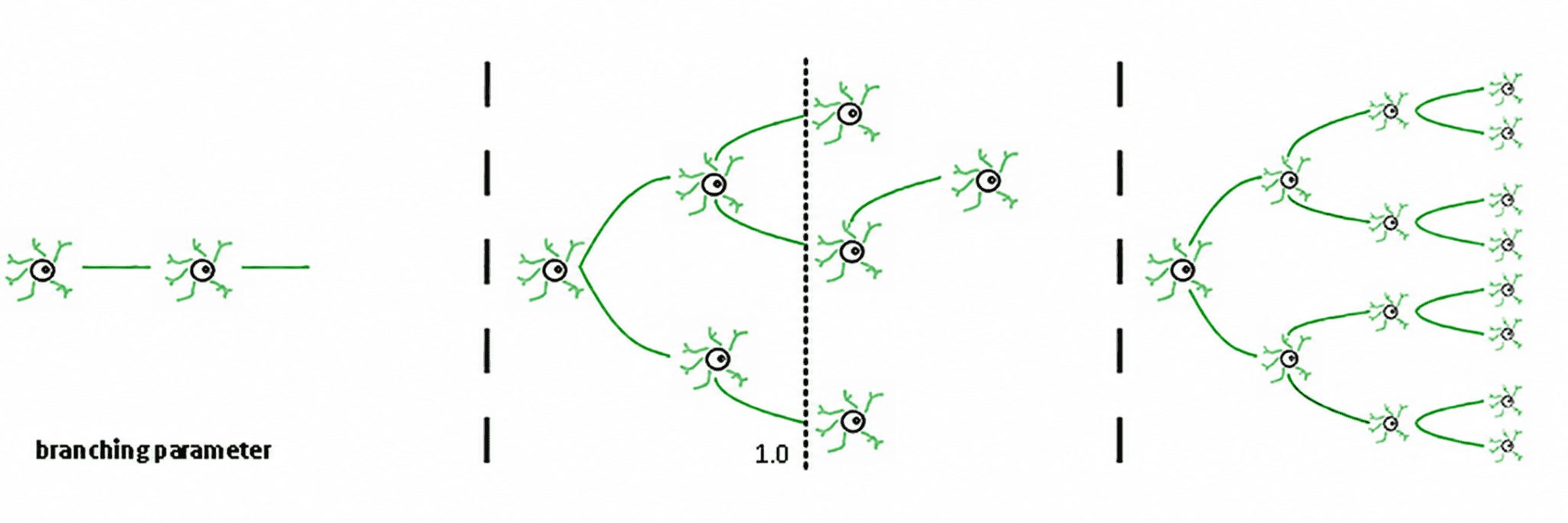
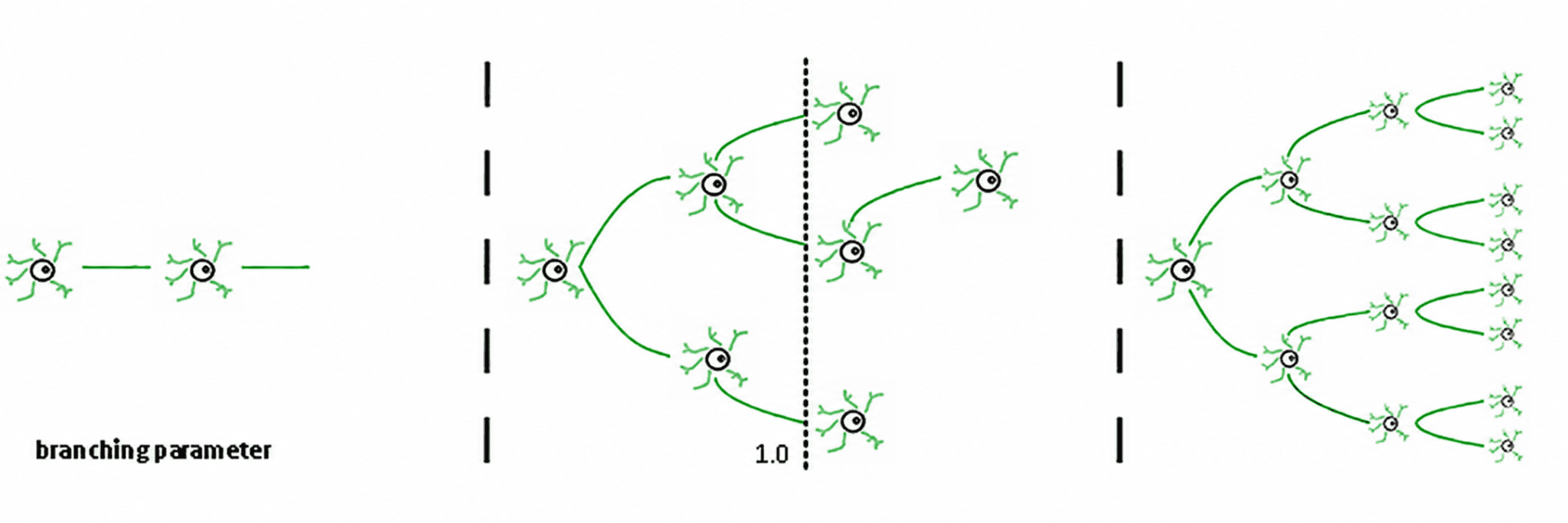
De esto se derivan tres regímenes (de Carvalho y Prado, 2000; Haldeman y Beggs, 2005):
Subcrítico (σ < 1): la actividad decae; el sistema "olvida" rápidamente la perturbación. Es estable, pero tiene poca memoria y no es muy expresivo.
Supercrítico (σ > 1): la actividad se dispara en cascadas. Esta es la característica distintiva de regímenes patológicos como las crisis epilépticas (Hsu et al., 2008; Hagemann et al., 2021).
Crítico (σ ≈ 1): cada pico, en promedio, genera otro pico. La actividad reverbera, las avalanchas neuronales obedecen leyes de potencia y el sistema mantiene una memoria estructurada de la entrada.
La belleza de σ reside en que es un único número que resume el régimen dinámico global. Sin embargo, su medición no es tan sencilla. Al aplicarse a registros corticales in vivo, la medición revela que la corteza no opera exactamente en σ = 1, sino ligeramente por debajo, en un régimen que los autores denominan reverberante (Wilting et al., 2018). La diferencia es importante: estar exactamente en σ = 1 sería como pedalear en bicicleta sobre una cuerda floja; estar ligeramente por debajo permite un ajuste rápido a las exigencias de la tarea sin riesgo de un colapso.
La criticidad en las redes neuronales artificiales: del borde del caos a la computación de reservorio
Bertschinger y Natschläger (2004) demostraron que las redes recurrentes aleatorias de umbral alcanzan su capacidad computacional máxima en tareas de procesamiento temporal precisamente en la transición orden-caos.
Boedecker et al. (2012) extendieron el análisis a redes de estado de eco dentro del paradigma de computación de reservorio, confirmando que la capacidad de transferencia de información y la memoria activa se maximizan en el borde del caos.
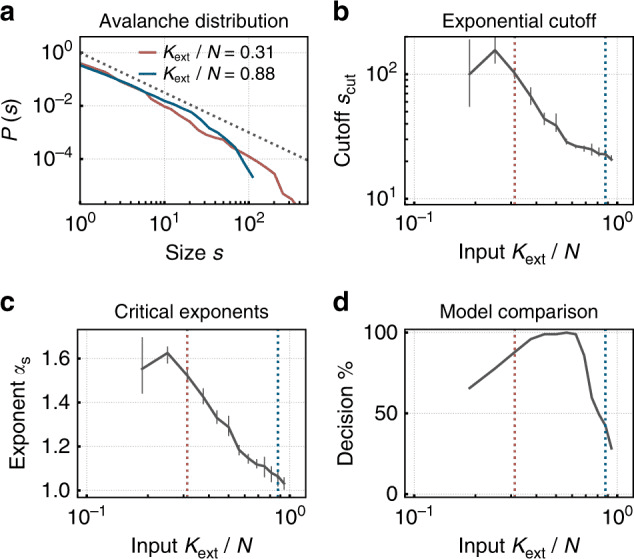
Figura 3. Una red neuromórfica de picos con plasticidad sináptica se autoorganiza hacia la criticidad bajo una entrada externa baja, exhibiendo distribuciones de tamaño de avalancha de ley de potencias, el sello distintivo del estado crítico tanto en redes neuronales biológicas como artificiales. Bajo una entrada mayor, la red cambia a un régimen subcrítico con distribuciones truncadas. Reproducido de Cramer et al. (2020), Nature Communications, 11, 2853. CC BY 4.0.
En el lenguaje de las redes neuronales artificiales, el parámetro de medición se denomina radio espectral. Cuando este supera 1, las trayectorias divergen exponencialmente (caos); cuando es muy inferior a 1, la red colapsa al punto fijo y pierde memoria. En este contexto, un radio espectral cercano a 1 es el equivalente formal del valor biológico σ ≈ 1 (Magnasco, 2022; Morales et al., 2023). En las redes neuronales de impulsos, la relación de ramificación se puede medir con métodos prácticamente idénticos a los utilizados en cultivos neuronales (Cramer et al., 2020; Zeraati et al., 2024).
¿Por qué la criticidad cerebral maximiza el cálculo neuronal?
Operar cerca de σ ≈ 1 proporciona cuatro ventajas que son fundamentales tanto para la hipótesis del cerebro crítico como para el diseño de sistemas de IA inspirados en el cerebro:
Rango dinámico máximo. Shew et al. (2009) demostraron que el rango de intensidades de entrada que la corteza puede discriminar es máximo cuando el equilibrio excitación-inhibición coloca la red en un punto crítico.
Capacidad de información maximizada. La entropía de los patrones de avalancha y la información mutua entre la entrada y la salida alcanzan su valor máximo en σ ≈ 1 (Shew et al., 2011).
Memoria de desvanecimiento óptima. En el régimen crítico, la perturbación se mantiene el tiempo justo para influir en el procesamiento sin contaminar el futuro lejano; es el punto óptimo entre la estabilidad y la integración temporal (Boedecker et al., 2012).
La complejidad como medida unificadora. Timme et al. (2016) demostraron que la complejidad neuronal se maximiza precisamente en el punto crítico, vinculando la criticidad con las teorías formales de la conciencia y el procesamiento.
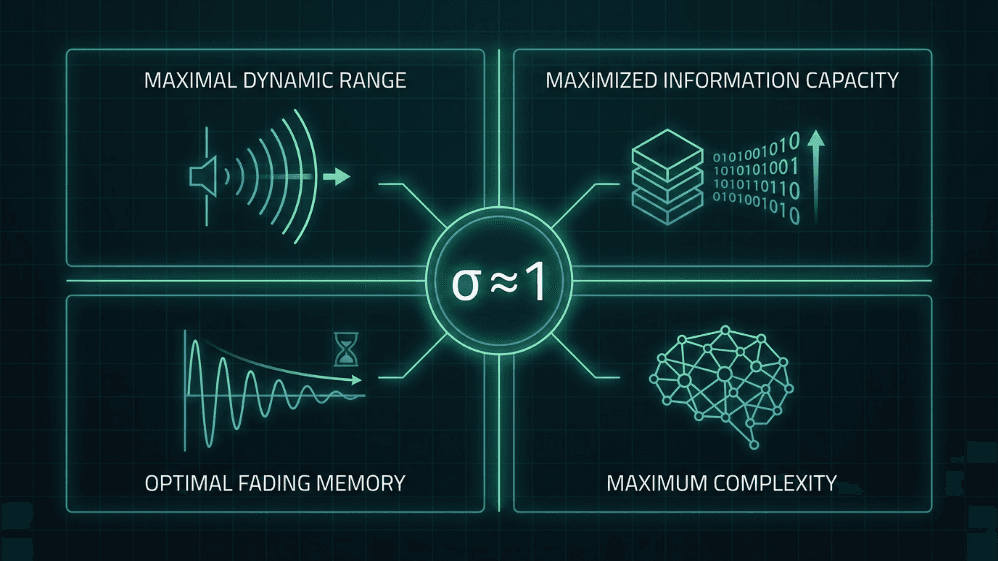
Figura 4. Cuatro ventajas computacionales de operar cerca de la relación de ramificación crítica (σ ≈ 1). En la criticidad, las redes neuronales alcanzan el máximo rango dinámico, la máxima capacidad de información, la memoria de desvanecimiento óptima y la máxima complejidad; propiedades que son fundamentales tanto para la hipótesis del cerebro crítico como para el diseño de IA inspirado en el cerebro.
El cerebro no siempre funciona a σ = 1
Esto no implica que el cerebro siempre opere a σ = 1. La evidencia sugiere más bien un régimen ligeramente subcrítico y modulable: durante tareas exigentes, la red se acerca a la criticidad; durante el sueño profundo, se aleja; y los estados patológicos (epilepsia, anestesia profunda, ciertas afecciones psiquiátricas) se asocian con desviaciones medibles de este rango operativo (Meisel et al., 2017; Zimmern, 2020). La relación de ramificación se está convirtiendo en un biomarcador dinámico del estado funcional del sistema nervioso.
Por qué utilizamos la relación de ramificación en Neuraxon: Diseño de IA bioinspirada al borde del caos
Neuraxon es un sistema bioinspirado que adopta principios dinámicos de la corteza como restricciones de diseño. La proporción de ramificación es una de las más importantes, y la utilizamos por cuatro razones:
Como invariante operacional en tiempo real para la estabilidad de la red neuronal
En arquitecturas recurrentes o de picos profundos, el doble riesgo de colapso de la actividad (red silenciosa, gradientes evanescentes) y explosión descontrolada (saturación, gradientes explosivos) es estructural. La monitorización de σ en tiempo real nos proporciona un único escalar diagnóstico, independiente de la arquitectura concreta, que indica si el sistema está activo desde el punto de vista computacional.
Como objetivo de autorregulación bioinspirada a través de la criticidad autoorganizada
La red se autoorganiza hacia la criticidad sin necesidad de un ajuste centralizado, replicando el principio de criticidad autoorganizada (Bornholdt y Röhl, 2003; Levina et al., 2007). Esto reduce drásticamente la sensibilidad a los hiperparámetros y dota al sistema de robustez frente a cambios en la distribución. Como exploramos en el volumen 7 de NIA sobre vida artificial y ecosistemas digitales, así es precisamente como surge la complejidad emergente a partir de reglas locales sin control centralizado.
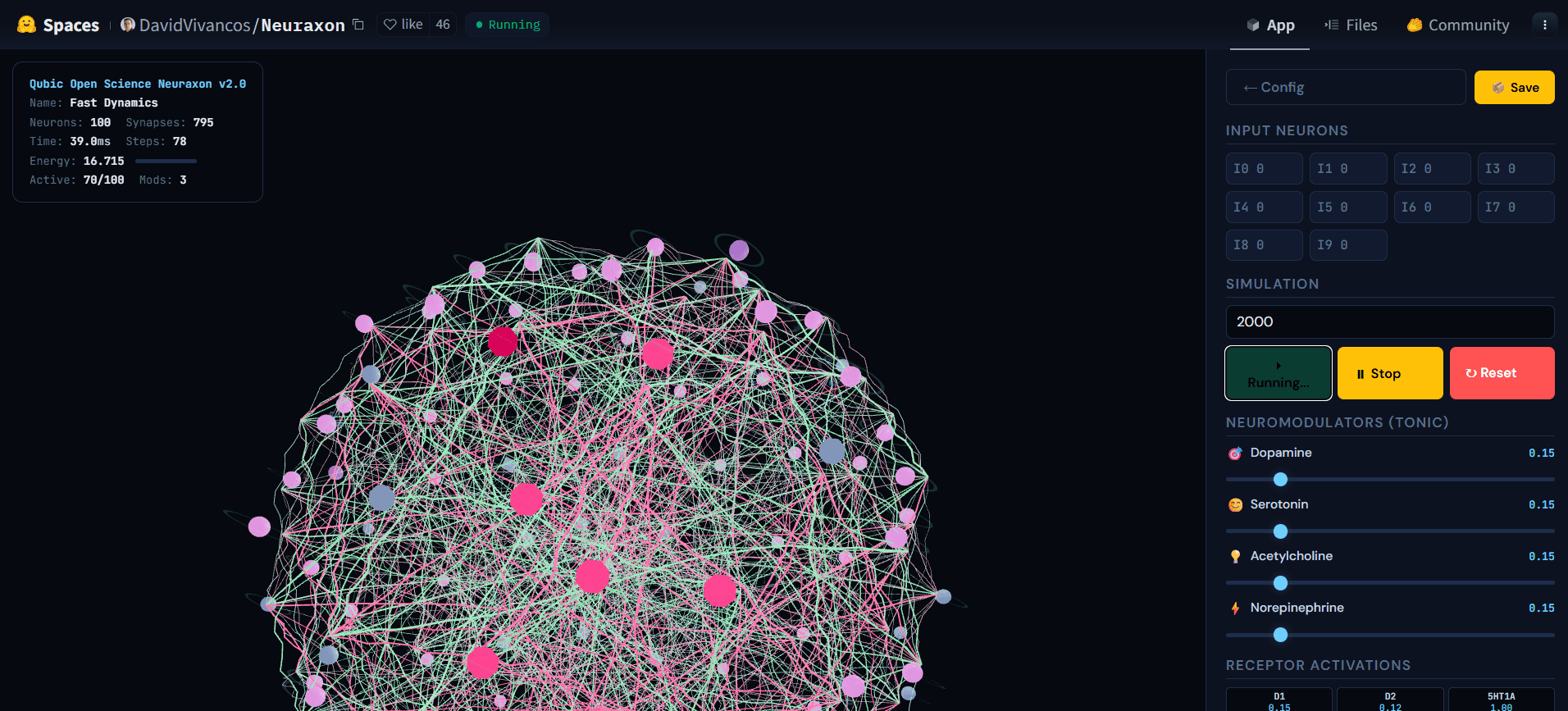
Figura 5. Red 3D de Neuraxon durante la simulación activa, que muestra la actividad en cascada a través de neuronas de estado ternario. Los nodos brillantemente activos (rosa) propagan señales a través de conexiones excitatorias (verde) e inhibitorias (rosa), mientras que otras neuronas permanecen en reposo (gris), ilustrando un régimen de reverberación cerca de la relación de ramificación crítica (σ ≈ 1). Este estado equilibrado —ni silencioso ni explosivo— es hacia el que Neuraxon se autoorganiza utilizando principios de criticidad bioinspirados. Explore la demostración interactiva en huggingface.co/spaces/DavidVivancos/Neuraxon. Fuente: Equipo Científico de Qubic.
Como puente entre la observación neurocientífica y el diseño de IA
La relación de ramificación es una de las pocas magnitudes que se miden con el mismo formalismo en electrofisiología, resonancia magnética funcional y redes artificiales. Esto permite probar hipótesis bidireccionales: si una intervención mejora la criticidad biológica, podemos preguntarnos si la misma intervención —traducida a la arquitectura artificial— mejora el cálculo del modelo, y viceversa. Este principio es fundamental para el marco de la neuromodulación y los mecanismos de activación astrocítica que hemos desarrollado en volúmenes anteriores de esta academia.
Como criterio funcional, no estético, para la IA inspirada en el cerebro
La criticidad es una restricción operativa con consecuencias empíricas. Operar cerca del régimen de reverberación mejora —según nuestras evaluaciones internas y publicaciones— la capacidad de generalización, la estabilidad ante perturbaciones de entrada, la riqueza representacional y la coherencia temporal del razonamiento. Estos efectos coinciden cualitativamente con los descritos en la literatura biológica (Cocchi et al., 2017) y artificial (Cramer et al., 2020; Morales et al., 2023).
La relación de ramificación: de la física estadística a la arquitectura de IA inspirada en el cerebro.
La relación de ramificación es una de esas rarezas conceptuales: lo suficientemente simple como para reducirla a una sola fórmula, pero lo suficientemente profunda como para conectar la física estadística, la neurociencia, la IA y el diseño de sistemas. Para el cerebro biológico, σ ≈ 1 parece ser el régimen donde emerge la combinación óptima de sensibilidad, memoria, expresividad y robustez. Para las redes artificiales, esta misma frontera —rebautizada como el límite del caos— predice la máxima capacidad computacional.
Para Neuraxon, se trata de un principio rector del diseño bioinspirado: una métrica auditable, autorregulada y biológicamente significativa que nos ayuda a mantener el sistema vivo, en el sentido más amplio de la palabra.
Referencias
Beggs, J. M., & Plenz, D. (2003). Avalanchas neuronales en circuitos neocorticales. The Journal of Neuroscience, 23(35), 11167–11177. https://doi.org/10.1523/JNEUROSCI.23-35-11167.2003
Bertschinger, N., & Natschläger, T. (2004). Computación en tiempo real al borde del caos en redes neuronales recurrentes. Neural Computation, 16(7), 1413–1436. https://doi.org/10.1162/089976604323057443
Boedecker, J., Obst, O., Lizier, J. T., Mayer, N. M., & Asada, M. (2012). Procesamiento de información en redes de estado de eco al borde del caos. Theory in Biosciences, 131(3), 205–213. https://doi.org/10.1007/s12064-011-0146-8
Bornholdt, S., & Röhl, T. (2003). Redes neuronales críticas autoorganizadas. Physical Review E, 67(6), 066118. https://doi.org/10.1103/PhysRevE.67.066118
Cocchi, L., Gollo, L. L., Zalesky, A., & Breakspear, M. (2017). Criticidad en el cerebro: Una síntesis de neurobiología, modelos y cognición. Progress in Neurobiology, 158, 132–152. https://doi.org/10.1016/j.pneurobio.2017.07.002
Cramer, B., Stöckel, D., Kreft, M., Wibral, M., Schemmel, J., Meier, K., & Priesemann, V. (2020). Control de criticidad y computación en redes neuromórficas de picos con plasticidad. Nature Communications, 11, 2853. https://doi.org/10.1038/s41467-020-16548-3
de Carvalho, J. X., & Prado, C. P. C. (2000). Criticidad autoorganizada en el modelo de Olami-Feder-Christensen. Physical Review Letters, 84(17), 4006–4009. https://doi.org/10.1103/PhysRevLett.84.4006
Derrida, B., & Pomeau, Y. (1986). Redes aleatorias de autómatas: Una aproximación recocida simple. Europhysics Letters, 1(2), 45–49. https://doi.org/10.1209/0295-5075/1/2/001
Hagemann, A., Wilting, J., Samimizad, B., Mormann, F., & Priesemann, V. (2021). Evaluación de la criticidad en la actividad de neuronas individuales pre-convulsivas de la corteza epiléptica humana. PLOS Computational Biology, 17(3), e1008773. https://doi.org/10.1371/journal.pcbi.1008773
Haldeman, C., & Beggs, J. M. (2005). La ramificación crítica captura la actividad en redes neuronales vivas y maximiza el número de estados metaestables. Physical Review Letters, 94(5), 058101. https://doi.org/10.1103/PhysRevLett.94.058101
Hsu, D., Chen, W., Hsu, M., & Beggs, J. M. (2008). Una hipótesis abierta: ¿Es la epilepsia aprendida y se puede desaprender? Epilepsy & Behavior, 13(3), 511–522. https://doi.org/10.1016/j.yebeh.2008.05.007
Langton, C. G. (1990). Computación al borde del caos: transiciones de fase y computación emergente. Physica D: Nonlinear Phenomena, 42(1–3), 12–37. https://doi.org/10.1016/0167-2789(90)90064-V
Levina, A., Herrmann, J. M., & Geisel, T. (2007). Sinapsis dinámicas que provocan criticidad autoorganizada en redes neuronales. Nature Physics, 3(12), 857–860. https://doi.org/10.1038/nphys758
Magnasco, M. O. (2022). Robustez y flexibilidad de la función neuronal a través de la criticidad dinámica. Entropy, 24(5), 591. https://doi.org/10.3390/e24050591
Meisel, C., Klaus, A., Vyazovskiy, V. V., & Plenz, D. (2017). La interacción entre las correlaciones temporales de largo y corto alcance moldea la dinámica cortical en diferentes estados de vigilia. The Journal of Neuroscience, 37(42), 10114–10124. https://doi.org/10.1523/JNEUROSCI.0448-17.2017
Morales, G. B., di Santo, S., & Muñoz, M. A. (2023). Desvelando la dinámica intrínseca de las redes neuronales biológicas y artificiales: De la criticidad a las representaciones óptimas. Frontiers in Complex Systems, 1, 1276338. https://doi.org/10.3389/fcpxs.2023.1276338
Plenz, D., Ribeiro, T. L., Miller, S. R., Kells, P. A., Vakili, A., & Capek, E. L. (2021). Criticidad autoorganizada en el cerebro. Frontiers in Physics, 9, 639389. https://doi.org/10.3389/fphy.2021.639389
Shew, W. L., Yang, H., Petermann, T., Roy, R., & Plenz, D. (2009). Las avalanchas neuronales implican un rango dinámico máximo en las redes corticales en el punto crítico. The Journal of Neuroscience, 29(49), 15595–15600. https://doi.org/10.1523/JNEUROSCI.3864-09.2009
Shew, W. L., Yang, H., Yu, S., Roy, R., & Plenz, D. (2011). La capacidad y transmisión de información se maximizan en redes corticales equilibradas con avalanchas neuronales. The Journal of Neuroscience, 31(1), 55–63. https://doi.org/10.1523/JNEUROSCI.4637-10.2011
Spitzner, F. P., Dehning, J., Wilting, J., Hagemann, A., Neto, J. P., Zierenberg, J., & Priesemann, V. (2021). MR. Estimator, una herramienta para determinar escalas de tiempo intrínsecas a partir de la actividad de picos submuestreada. PLOS ONE, 16(4), e0249447. https://doi.org/10.1371/journal.pone.0249447
Timme, N. M., Marshall, N. J., Bennett, N., Ripp, M., Lautzenhiser, E., & Beggs, J. M. (2016). La criticidad maximiza la complejidad en el tejido neural. Frontiers in Physiology, 7, 425. https://doi.org/10.3389/fphys.2016.00425
Turrigiano, G. G. (2008). La neurona autoajustable: Escalamiento sináptico de las sinapsis excitatorias. Cell, 135(3), 422–435. https://doi.org/10.1016/j.cell.2008.10.008
Wang, J., Cao, R., Brunton, B. W., Smith, R. E. W., Buckner, R. L., & Liu, T. T. (2025). Contribuciones genéticas a la criticidad cerebral y su relación con las funciones cognitivas humanas. Proceedings of the National Academy of Sciences, 122(26), e2417010122. https://doi.org/10.1073/pnas.2417010122
Wilting, J., Dehning, J., Pinheiro Neto, J., Rudelt, L., Wibral, M., Zierenberg, J., & Priesemann, V. (2018). Operar en un régimen de reverberación permite una rápida sintonización de los estados de la red a los requisitos de la tarea. Frontiers in Systems Neuroscience, 12, 55. https://doi.org/10.3389/fnsys.2018.00055
Wilting, J., & Priesemann, V. (2018). Inferencia de estados dinámicos colectivos a partir de sistemas poco observados. Nature Communications, 9, 2325. https://doi.org/10.1038/s41467-018-04725-4
Wilting, J., & Priesemann, V. (2019). 25 años de análisis crítico en neurociencia: resultados establecidos, controversias abiertas, conceptos novedosos. Current Opinion in Neurobiology, 58, 105–111. https://doi.org/10.1016/j.conb.2019.08.002
Yu, C. (2022). Hacia un análisis unificado de la hipótesis de criticidad cerebral: Revisión de varias herramientas disponibles. Frontiers in Neural Circuits, 16, 911245. https://doi.org/10.3389/fncir.2022.911245
Zeraati, R., Engel, T. A., & Levina, A. (2024). Estimación de escalas temporales intrínsecas y criticidad a partir de registros neuronales: Métodos y dificultades. Current Opinion in Neurobiology, 86, 102871. https://doi.org/10.1016/j.conb.2024.102871
Zimmern, V. (2020). Por qué la criticidad cerebral es clínicamente relevante: Una revisión exploratoria. Frontiers in Neural Circuits, 14, 54. https://doi.org/10.3389/fncir.2020.00054
Explora la Academia de Inteligencia Neuroaxon completa
Este es el volumen 8 de #Neuraxon Inteligencia #academy del #Qubic Equipo Científico. Si te acabas de unir, explora la serie completa para comprender a fondo la ciencia detrás de Neuraxon, #aigarth y el enfoque de Qubic hacia la #decentralized inteligencia artificial inspirada en el cerebro:
NIA Vol. 1: Por qué la inteligencia no se calcula en pasos, sino en el tiempo — Explora por qué la inteligencia biológica opera en tiempo continuo en lugar de pasos computacionales discretos como los modelos LLM tradicionales.
NIA Vol. 2: Dinámica ternaria como modelo de inteligencia viva — Explica la dinámica ternaria y por qué la lógica de tres estados (excitatorio, neutro, inhibitorio) es importante para modelar sistemas vivos.
NIA Vol. 3: Neuromodulación e IA inspirada en el cerebro: abarca la neuromodulación y cómo la señalización química del cerebro (dopamina, serotonina, acetilcolina, norepinefrina) inspira la arquitectura de Neuraxon.
NIA Vol. 4: Redes neuronales en IA y neurociencia: una comparación profunda de redes neuronales biológicas, redes neuronales artificiales y el enfoque de tercera vía de Neuraxon.
NIA Vol. 5: Astrocitos e IA inspirada en el cerebro: cómo la activación astrocítica transforma la plasticidad de la red neuronal a través del marco AGMP en Neuraxon.
NIA Vol. 6: Máquinas conscientes frente a organismos inteligentes: la consciencia de la IA explicada — Explora la consciencia de la IA a través de la lente de la teoría del espacio de trabajo global, la teoría de la información integrada y la codificación predictiva.
NIA Vol. 7: El juego de la vida de Conway, la vida artificial y los ecosistemas digitales: la ciencia detrás del enfoque de Qubic, Aigarth y Neuraxon sobre la complejidad emergente y la criticidad autoorganizada en la IA descentralizada.
Qubic es una red descentralizada de código abierto para tecnología experimental. Para obtener más información, visita qubic.org. Únete a la conversación en X, Discord y Telegram.